Taxonomia de la Memoria
Durante gran parte del siglo XX, los cientificos trataron la memoria como un sistema unitario: un almacen donde se guardaban las experiencias. Hoy sabemos que esa idea es profundamente incorrecta. La memoria es un conjunto de sistemas independientes, cada uno con su propia neuroanatomia, sus propias reglas de codificacion y sus propios mecanismos de recuperacion. Danada una estructura cerebral, se pierde un tipo de memoria pero los demas permanecen intactos. Esta disociacion multiple es la evidencia mas poderosa de que los sistemas son genuinamente distintos.
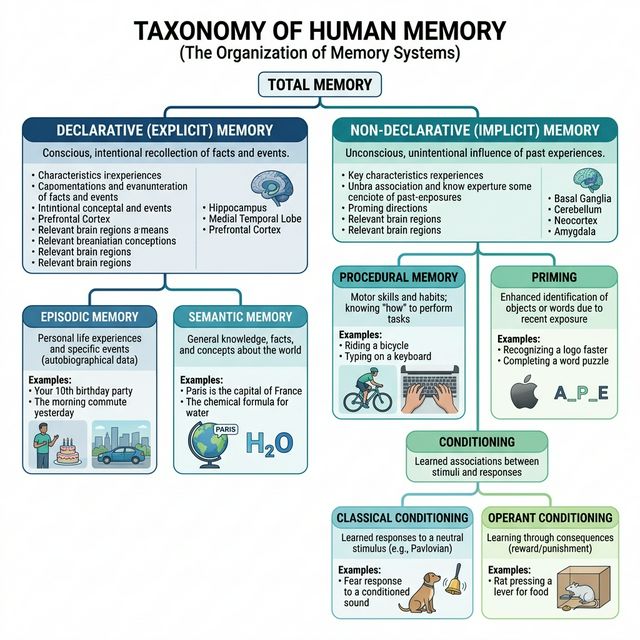
Memoria declarativa (explicita)
La memoria declarativa es aquella que podemos expresar verbalmente, "declarar" de forma consciente. Depende criticamente del hipocampo y de las cortezas temporales mediales. Se subdivide en dos grandes ramas:
- Memoria episodica: Es el sistema que nos permite revivir experiencias personales situadas en un tiempo y lugar concretos. Cuando recuerdas tu primer dia de universidad, estas accediendo a una memoria episodica: puedes visualizar el edificio, sentir la ansiedad, recordar con quien hablaste. Endel Tulving, quien propuso esta distincion en 1972, la describio como "viaje mental en el tiempo". Las memorias episodicas son ricas en contexto, estan cargadas de emocion y son altamente susceptibles al olvido. Se codifican en el hipocampo y gradualmente se transfieren a la neocorteza durante la consolidacion. Un dato fascinante: los taxistas de Londres, que navegan miles de calles, muestran un hipocampo posterior significativamente mas grande que el promedio, lo que refleja la plasticidad de esta estructura ante la demanda episodica-espacial.
- Memoria semantica: Almacena conocimiento general sobre el mundo, independiente del contexto en que fue aprendido. Sabes que Paris es la capital de Francia, que el agua hierve a 100 grados y que los mamiferos son de sangre caliente, pero probablemente no recuerdas exactamente cuando ni donde aprendiste cada uno de estos datos. La memoria semantica esta distribuida ampliamente en la neocorteza, particularmente en los lobulos temporales laterales e inferiores. A diferencia de la episodica, la memoria semantica es mas resistente al olvido y puede sobrevivir incluso a un dano hipocampal considerable, siempre que haya habido tiempo suficiente para la consolidacion cortical.
Memoria no declarativa (implicita)
La memoria no declarativa abarca formas de aprendizaje que no requieren acceso consciente y que se expresan a traves de la conducta, no de la narracion verbal. Incluye varios subsistemas distintos:
- Memoria procedimental: Es el saber "como" hacer algo: andar en bicicleta, tocar el piano, escribir a maquina. Depende de los ganglios basales (especialmente el cuerpo estriado) y del cerebelo. Se adquiere lentamente a traves de la practica repetida, pero una vez consolidada es extraordinariamente duradera. Un pianista profesional puede dejar de tocar durante anos y recuperar la habilidad mucho mas rapido que alguien que la aprende por primera vez. Esta forma de memoria fue la que se preservo intacta en el paciente H.M., lo que demostro de forma concluyente que el hipocampo no es necesario para todo tipo de aprendizaje.
- Priming (facilitacion): Se refiere a la influencia que tiene la exposicion previa a un estimulo sobre el procesamiento posterior del mismo estimulo o de estimulos relacionados. Si lees la palabra "DOCTOR" y luego te muestran la palabra incompleta "ENF_____", es mas probable que la completes como "ENFERMERA" que como "ENFADAR". El priming ocurre en las cortezas sensoriales y de asociacion, y es independiente del hipocampo. Funciona incluso cuando la persona no recuerda conscientemente haber visto el estimulo previo, lo que demuestra su naturaleza implicita.
- Condicionamiento clasico: El aprendizaje asociativo descubierto por Pavlov. Un estimulo neutro (campana) se asocia con un estimulo biologicamente significativo (comida) hasta que el estimulo neutro por si solo produce la respuesta (salivacion). El condicionamiento de respuestas emocionales depende de la amigdala, mientras que el condicionamiento de respuestas motoras (como el parpadeo condicionado) depende del cerebelo. Este tipo de memoria es filogeneticamente antiguo y se observa desde invertebrados hasta humanos.
Memoria de trabajo: el modelo de Baddeley
La memoria de trabajo no es simplemente "memoria a corto plazo". Es un sistema activo de manipulacion de informacion que nos permite mantener y operar sobre datos en tiempo real. Alan Baddeley propuso en 1974 un modelo influyente que incluye cuatro componentes:
- Bucle fonologico: Mantiene informacion verbal mediante repeticion subvocal. Es lo que usas cuando repites un numero de telefono mentalmente hasta que lo marcas. Tiene una capacidad limitada (aproximadamente 2 segundos de habla) y depende de areas del lobulo frontal inferior izquierdo y del lobulo parietal inferior.
- Agenda visuoespacial: Mantiene imagenes mentales y representaciones espaciales. Es lo que usas cuando visualizas el recorrido de tu casa al trabajo o cuando rotas mentalmente un objeto. Involucra cortezas parietales y occipitales derechas.
- Buffer episodico: Anadido al modelo en 2000, es un espacio de integracion multimodal que combina informacion del bucle fonologico, la agenda visuoespacial y la memoria a largo plazo en representaciones coherentes. Es esencial para la comprension de narraciones y la planificacion.
- Ejecutivo central: El director de orquesta. Controla la atencion, selecciona estrategias y coordina los otros subsistemas. Se asocia con la corteza prefrontal dorsolateral. Su disfuncion se manifiesta en trastornos como el TDAH y en el deterioro cognitivo asociado al envejecimiento.
La capacidad de la memoria de trabajo predice fuertemente el rendimiento academico, la inteligencia fluida y la capacidad de razonamiento. No es una cifra fija: puede entrenarse parcialmente, y varia segun el estado de alerta, el estres y la motivacion.
El Hipocampo y la Consolidacion
Codificacion: del estimulo a la huella
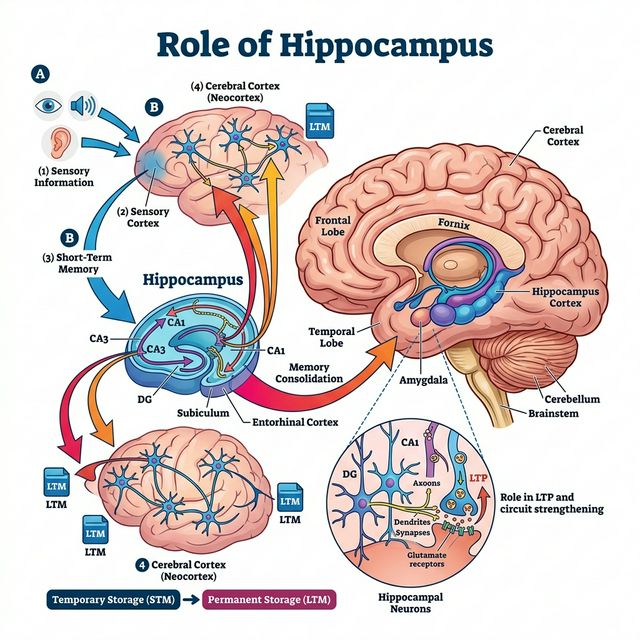
La codificacion es el primer paso en la formacion de una memoria. Cuando experimentas un evento, multiples areas corticales procesan simultaneamente los diferentes aspectos de la experiencia: las cortezas visuales procesan las imagenes, las cortezas auditivas los sonidos, las cortezas somatosensoriales las sensaciones tactiles, y la amigdala evalua el significado emocional. El hipocampo actua como un "centro de enlace" que conecta estas representaciones distribuidas en un patron unificado, creando lo que los neurocientificos llaman una "huella de memoria" o engrama.
La calidad de la codificacion depende criticamente de la atencion y de la profundidad del procesamiento. Fergus Craik y Robert Lockhart demostraron en 1972 que cuanto mas profundamente procesamos la informacion (analizando su significado, conectandola con conocimiento previo, elaborandola), mejor se retiene. Leer una palabra y juzgar si rima con otra (procesamiento superficial) produce memorias mucho mas debiles que leer la misma palabra y juzgar si encaja en una oracion (procesamiento profundo). Este efecto de niveles de procesamiento tiene profundas implicaciones educativas.
Consolidacion sinaptica: las primeras horas
Inmediatamente despues de la codificacion, la huella de memoria es extremadamente fragil. Durante las primeras horas, ocurre un proceso llamado consolidacion sinaptica, que involucra cambios moleculares en las sinapsis del hipocampo. La activacion de receptores NMDA permite la entrada de calcio, que desencadena cascadas de senalizacion intracelular (incluyendo CREB, BDNF y sintesis de nuevas proteinas) que fortalecen las conexiones sinapticas. Si este proceso se interrumpe -- por ejemplo, con inhibidores de la sintesis de proteinas o por un traumatismo craneoencefalico -- la memoria no se consolida y se pierde permanentemente. Esta es la base de la amnesia retrograda post-traumatica: los eventos inmediatamente anteriores al trauma no tuvieron tiempo de consolidarse.
Consolidacion de sistemas: meses y anos
Existe un segundo nivel de consolidacion, mucho mas lento, llamado consolidacion de sistemas. Segun la teoria estandar de consolidacion, propuesta por Larry Squire y otros, las memorias declarativas inicialmente dependen del hipocampo pero gradualmente se transfieren a la neocorteza durante un periodo que puede durar meses o incluso anos. Este proceso explica por que pacientes con dano hipocampal pueden recordar eventos lejanos (ya consolidados en la corteza) pero no eventos recientes (aun dependientes del hipocampo).
Sin embargo, la teoria de las multiples huellas de Morris Moscovitch sugiere una variacion importante: las memorias episodicas ricas en detalle contextual siempre requieren algo de participacion hipocampal, incluso despues de anos. Solo las memorias que se han "semantizado" -- transformandose en conocimiento factual desprovisto de contexto -- se independizan completamente del hipocampo. El debate entre estas dos teorias sigue activo.
Reactivacion hipocampal durante el sueno
El mecanismo clave de la consolidacion de sistemas es la reactivacion hipocampal durante el sueno, especialmente durante el sueno de ondas lentas (NREM fase 3). Las neuronas del hipocampo que estuvieron activas durante el aprendizaje diurno se reactivan espontaneamente durante el sueno, pero a una velocidad mucho mayor (hasta 20 veces mas rapido). Este "replay comprimido" se coordina con oscilaciones de la neocorteza (ondas lentas y husos de sueno) para transferir gradualmente la informacion del hipocampo a redes corticales de almacenamiento a largo plazo.
Experimentos elegantes han demostrado este fenomeno en ratas: John O'Keefe y sus colegas registraron neuronas de lugar en el hipocampo mientras las ratas navegaban un laberinto. Durante el sueno posterior, las mismas secuencias de disparo neuronal se repetian en el mismo orden, como si las ratas "sonaran" con el recorrido del laberinto. En humanos, estudios con fMRI y EEG han mostrado patrones similares de reactivacion.
La importancia practica es enorme: dormir despues de estudiar mejora significativamente la retencion. Incluso una siesta de 90 minutos (suficiente para un ciclo de sueno completo con fase NREM profunda) puede mejorar el rendimiento en tareas de memoria declarativa entre un 15% y un 40%.
Olvido, Interferencia y Memoria Emocional
La curva del olvido de Ebbinghaus
En 1885, Hermann Ebbinghaus realizo el primer estudio cientifico del olvido, utilizandose a si mismo como unico sujeto experimental. Memorizo listas de silabas sin sentido (como "ZOL", "DAK", "BUP") y luego midio cuanto retenia en intervalos que iban desde 20 minutos hasta 31 dias. Su descubrimiento fue tan elegante como perturbador: el olvido sigue una funcion exponencial negativa. En los primeros 20 minutos se pierde aproximadamente el 40% de lo aprendido. Despues de una hora, se ha olvidado el 56%. Despues de un dia, el 66%. Despues de una semana, el 75%. La curva se aplana progresivamente: lo que sobrevive a la primera semana tiene buenas probabilidades de perdurar.
Esta curva tiene una implicacion practica inmediata: la repeticion espaciada. Si revisas el material justo antes de olvidarlo, cada repeticion aplana la curva un poco mas, extendiendo el intervalo de retencion. Este principio es la base de sistemas de estudio como las tarjetas Anki y la tecnica de Leitner. Ebbinghaus tambien descubrio el "efecto de ahorro": incluso cuando crees haber olvidado completamente algo, reaprender ese material lleva menos tiempo que aprenderlo desde cero, lo que indica que alguna huella permanece.
Interferencia proactiva y retroactiva
El olvido no ocurre solo por el paso del tiempo. Mucho de lo que perdemos se debe a la interferencia entre memorias competidoras. Existen dos formas principales:
- Interferencia proactiva: Las memorias antiguas dificultan el aprendizaje de informacion nueva. Si has conducido autos con cambio manual durante anos y cambias a uno automatico, tu mano seguira buscando la palanca de cambios. Las viejas asociaciones motoras interfieren con la nueva conducta. En el laboratorio, si aprendes la lista A y luego la lista B, tu recuerdo de la lista B sera peor que si solo hubieras aprendido la lista B, porque la lista A "contamina" la recuperacion.
- Interferencia retroactiva: La informacion nueva debilita las memorias antiguas. Si aprendes vocabulario en frances y luego aprendes vocabulario en italiano, tu recuerdo del frances empeorara. Este efecto es especialmente fuerte cuando los materiales son similares. La interferencia retroactiva explica por que estudiar para dos examenes el mismo dia produce peores resultados que separarlos: los contenidos del segundo examen interfieren retroactivamente con los del primero.
El debate entre las teorias del decaimiento (las memorias simplemente se debilitan con el tiempo) y la interferencia (las memorias se bloquean mutuamente) ha sido central en la psicologia de la memoria. La evidencia actual sugiere que ambos mecanismos operan simultaneamente, pero la interferencia es probablemente la causa predominante del olvido cotidiano.
Modulacion amigdalina de la memoria emocional
No todas las memorias se crean iguales. Los eventos emocionalmente intensos -- tanto positivos como negativos -- se recuerdan con mucha mayor vivacidad y durabilidad que los eventos neutros. Este fenomeno se debe a la modulacion que ejerce la amigdala sobre los procesos de consolidacion hipocampal.
El mecanismo funciona asi: cuando experimentas un evento emocionalmente significativo, la amigdala se activa y libera noradrenalina y cortisol (hormonas del estres). Estas sustancias actuan sobre el hipocampo y sobre las cortezas sensoriales, potenciando la consolidacion sinaptica y haciendo que la huella de memoria sea mas fuerte y mas detallada. James McGaugh, pionero en este campo, demostro que si se bloquean farmacologicamente los receptores beta-adrenergicos (con propranolol, por ejemplo), se elimina la ventaja mnemonica de los eventos emocionales sin afectar el recuerdo de eventos neutros.
Este mecanismo tiene sentido evolutivo: recordar donde encontraste comida o donde te ataco un depredador tiene valor de supervivencia. Pero en la patologia, el mismo mecanismo subyace al trastorno de estres postraumatico (TEPT), donde memorias traumaticas se consolidan con tal fuerza que se vuelven intrusivas e incontrolables.
Memorias flash (flashbulb memories)
Un tipo especial de memoria emocional son las llamadas memorias flash o flashbulb memories, descritas por Roger Brown y James Kulik en 1977. Son memorias extraordinariamente vividas de las circunstancias en las que te enteraste de un evento publico impactante. Muchas personas recuerdan exactamente donde estaban y que hacian cuando se enteraron de los atentados del 11 de septiembre de 2001, o cuando murio un ser querido.
Durante decadas se creyo que estas memorias eran fotograficamente precisas, como si la emocion "congelara" el momento. Sin embargo, estudios longitudinales (como los de Ulric Neisser) han demostrado que las memorias flash, aunque subjetivamente muy vividas y cargadas de certeza, contienen tantas distorsiones y errores como las memorias ordinarias. La emocion aumenta la confianza subjetiva en el recuerdo, pero no necesariamente su precision. Esta disociacion entre vivacidad y exactitud es uno de los hallazgos mas contraintuitivos de la ciencia de la memoria.
Reconsolidacion: las memorias se reescriben cada vez que se recuerdan
Quizas el descubrimiento mas revolucionario de las ultimas dos decadas en el campo de la memoria es la reconsolidacion. Hasta finales de los anos 90, se asumia que una vez consolidada, una memoria era permanente e inmutable. En el ano 2000, Karim Nader, Glenn Schafe y Joseph LeDoux demostraron que cuando una memoria consolidada se reactiva (se recuerda), entra temporalmente en un estado labil y debe volver a consolidarse -- un proceso que requiere nueva sintesis de proteinas, igual que la consolidacion original.
Esto significa que cada vez que recuerdas algo, ese recuerdo se "abre" bioquimicamente, puede ser modificado, y luego se "guarda" de nuevo. Si el proceso de reconsolidacion se interrumpe (farmacologicamente o mediante interferencia conductual), la memoria puede debilitarse o incluso borrarse. Las implicaciones terapeuticas son enormes: la reconsolidacion abre la puerta a tratamientos para el TEPT que podrian debilitar selectivamente memorias traumaticas, y explica por que los recuerdos se distorsionan progresivamente cada vez que los narramos.
El Paciente H.M.: Henry Molaison (1926-2008)
Henry Molaison sufrio epilepsia grave e intratable desde los 10 anos, tras un accidente en bicicleta. Para 1953, a los 27 anos, sus convulsiones eran tan frecuentes e incapacitantes que no podia trabajar ni llevar una vida normal. El neurocirujano William Becker Scoville propuso una solucion radical: extirpar bilateralmente las estructuras del lobulo temporal medial, incluyendo aproximadamente dos tercios del hipocampo, la corteza entorrinal, la corteza perirrinal y la amigdala. La cirugia fue un exito contra la epilepsia -- las convulsiones se redujeron dramaticamente. Pero el precio fue devastador.
Tras la operacion, Henry era incapaz de formar nuevas memorias declarativas. Podia recordar su infancia y eventos previos a la cirugia, pero cada persona que conocia, cada comida que comia, cada conversacion que tenia, desaparecia de su mente en cuestion de minutos. Leia la misma revista una y otra vez como si fuera nueva. No reconocia a la investigadora Brenda Milner que lo estudio durante decadas -- cada vez que ella entraba en la habitacion, Henry la saludaba como si fuera la primera vez que se encontraban.
Sin embargo, algo extraordinario emergio de los estudios: Henry podia aprender nuevas habilidades motoras. En la tarea del dibujo en espejo (trazar el contorno de una estrella mirando solo su reflejo), su rendimiento mejoraba sesion tras sesion, aunque cada dia negaba haber realizado la tarea antes. "Esto es curioso", decia, "pense que seria dificil, pero me esta saliendo bastante bien".
Doble disociacion entre sistemas de memoria: El caso de H.M. demostro de forma concluyente que la memoria declarativa (episodica y semantica) y la memoria procedimental son sistemas neuroanatomicamente distintos. El hipocampo y las estructuras temporales mediales son esenciales para la formacion de nuevas memorias declarativas, pero no para el aprendizaje procedimental.
Memoria procedimental = Ganglios basales + Cerebelo: Las habilidades motoras como el dibujo en espejo dependen del cuerpo estriado (parte de los ganglios basales) y del cerebelo, estructuras que estaban intactas en H.M. Estas regiones pueden formar nuevas memorias procedimentales sin ninguna participacion del hipocampo. Henry mejoraba en la tarea porque su sistema estriatal estaba aprendiendo, aunque su sistema hipocampal no podia registrar que la tarea habia ocurrido.
Consciencia y memoria son disociables: H.M. no tenia experiencia consciente de haber aprendido -- no recordaba las sesiones de practica. Pero su cerebro habia aprendido a nivel implicito. Esto demuestra que la consciencia del recuerdo (memoria explicita) y el aprendizaje conductual (memoria implicita) son procesos separados.
Impacto historico: El caso de H.M. transformo las neurociencias. Antes de el, no habia evidencia clara de que diferentes tipos de memoria dependieran de diferentes estructuras cerebrales. Su caso abrio todo el campo moderno de la neurociencia de la memoria y establecio al hipocampo como la estructura central de la memoria declarativa. Brenda Milner y Suzanne Corkin dedicaron sus carreras al estudio de H.M., produciendo mas de 100 publicaciones cientificas.