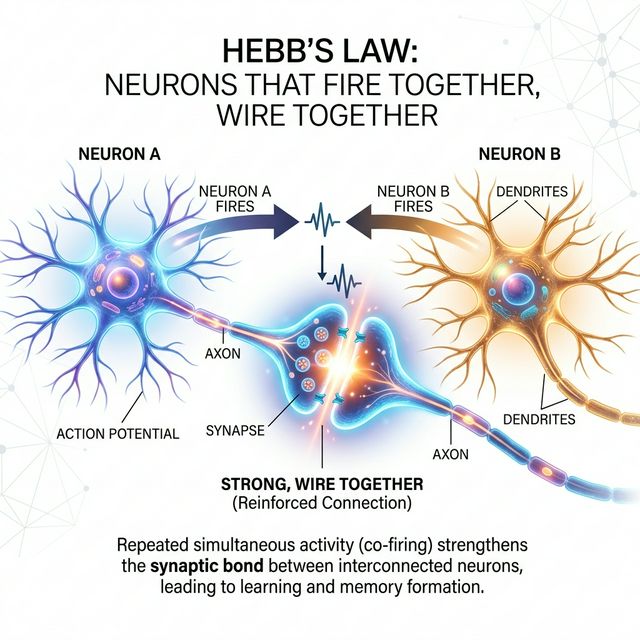
Ley de Hebb: Neuronas que disparan juntas, se conectan juntas
En 1949, el psicólogo canadiense Donald Olding Hebb publicó The Organization of Behavior, un libro que cambiaría para siempre nuestra comprensión de cómo el cerebro aprende. En él formuló un principio elegante y poderoso que hoy conocemos como la Ley de Hebb:
«Cuando un axón de la célula A está lo suficientemente cerca como para excitar a la célula B, y participa repetida o persistentemente en su disparo, algún proceso de crecimiento o cambio metabólico tiene lugar en una o ambas células, de tal manera que la eficiencia de A como una de las células que disparan a B, aumenta.»
Este principio se popularizó con la frase «neurons that fire together, wire together» (las neuronas que disparan juntas, se conectan juntas), acuñada por el neurocientífico Carla Shatz en 1992. Aunque es una simplificación, captura la esencia del aprendizaje asociativo a nivel celular.
El aprendizaje asociativo en acción
Imagina que estás aprendiendo a tocar el piano. Cada vez que ves una nota en la partitura (estímulo visual) y presionas la tecla correcta (respuesta motora), dos conjuntos de neuronas se activan simultáneamente: las de la corteza visual y las de la corteza motora. Según la Ley de Hebb, cada activación simultánea fortalece la conexión sináptica entre ambos grupos neuronales.
Al principio, la conexión es débil: necesitas pensar conscientemente dónde está cada tecla. Pero después de miles de repeticiones, la sinapsis se ha fortalecido tanto que la respuesta se vuelve automática: tus dedos «saben» dónde ir al ver la nota. Esto es exactamente lo que predice Hebb.
Propiedades clave del aprendizaje hebbiano
- Dependencia temporal: Las neuronas deben disparar dentro de una ventana temporal estrecha (milisegundos). Si la neurona A dispara mucho antes o después de B, la conexión no se fortalece — puede incluso debilitarse. Este refinamiento se conoce como STDP (Spike-Timing Dependent Plasticity).
- Especificidad: Solo se fortalecen las sinapsis específicas que participan en la actividad conjunta, no todas las sinapsis de la neurona.
- Cooperatividad: Se necesita un umbral mínimo de actividad simultánea para inducir el cambio — una sola sinapsis débil no basta.
- Asociatividad: Una entrada débil puede fortalecerse si se activa al mismo tiempo que una entrada fuerte, como un «efecto de arrastre» neuronal.
La genialidad de Hebb fue postular este mecanismo décadas antes de que se descubriera su base molecular. Hoy sabemos que el mecanismo que hace realidad la predicción de Hebb tiene un nombre: la Potenciación a Largo Plazo (LTP).
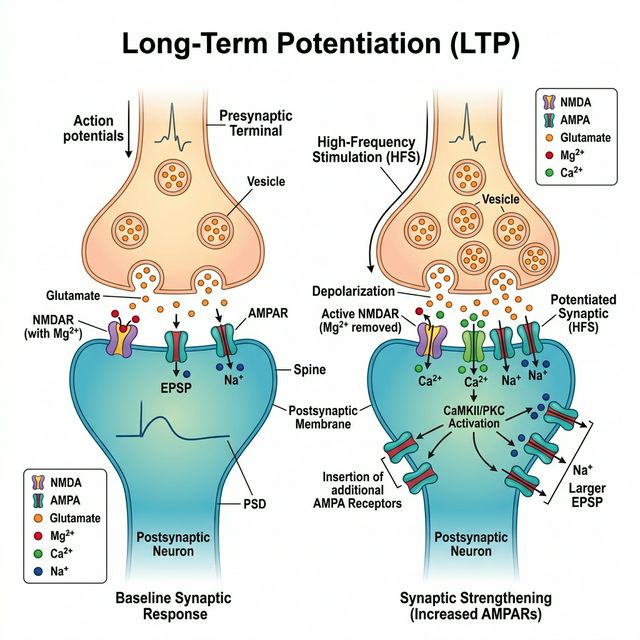
Mecanismos moleculares de la LTP
La Potenciación a Largo Plazo (LTP) fue descubierta en 1973 por Terje Lømo y Timothy Bliss en el hipocampo del conejo. Demostraron que una estimulación breve de alta frecuencia podía fortalecer una sinapsis durante horas, días o incluso semanas. La LTP es considerada el correlato celular de la memoria y el aprendizaje.
Paso 1: Transmisión basal — Los receptores AMPA
En condiciones normales, cuando la neurona presináptica libera glutamato (el neurotransmisor excitador principal del cerebro), este se une a los receptores AMPA (ácido α-amino-3-hidroxi-5-metil-4-isoxazolpropionico) en la membrana postsináptica. Los receptores AMPA son canales iónicos que permiten la entrada rápida de Na+ (sodio), generando una pequeña despolarización llamada PEPS (Potencial Excitador Postsináptico).
Esta despolarización por sí sola es insuficiente para inducir plasticidad. Es la transmisión sináptica «ordinaria», la señal basal que permite la comunicación neuronal momento a momento.
Paso 2: El detector de coincidencia — Los receptores NMDA
Los receptores NMDA (N-metil-D-aspartato) son los verdaderos protagonistas de la LTP. Son canales iónicos únicos porque funcionan como detectores de coincidencia: solo se abren cuando se cumplen dos condiciones simultáneas:
- Glutamato presente: La neurona presináptica debe estar activa (liberando glutamato).
- Despolarización suficiente: La neurona postsináptica debe estar ya parcialmente despolarizada (por la activación de múltiples receptores AMPA o por la llegada de otros estímulos).
¿Por qué se necesitan ambas condiciones? Porque en reposo, el canal del receptor NMDA está bloqueado por un ión de Magnesio (Mg2+) que actúa como un «tapón» dependiente de voltaje. Cuando la membrana se despolariza lo suficiente, el Mg2+ es expulsado electrostáticamente del canal, y si al mismo tiempo hay glutamato unido al receptor, el canal se abre.
Esta doble condición es la implementación molecular de la Ley de Hebb: el receptor NMDA solo se activa cuando ambas neuronas están activas simultáneamente.
Paso 3: La señal de calcio — El mensajero crítico
Cuando el receptor NMDA finalmente se abre, permite la entrada de un ión crucial: el Calcio (Ca2+). El calcio es mucho más que un simple ión — es un segundo mensajero intracelular que desencadena una cascada de eventos moleculares:
- Activación de CaMKII (proteína quinasa dependiente de calcio/calmodulina tipo II): Esta enzima es considerada la «molécula de la memoria». Una vez activada, puede autofosforilarse, manteniendo su actividad incluso después de que los niveles de calcio vuelvan a la normalidad. Es como un interruptor molecular que permanece «encendido».
- Activación de PKC (Proteína Quinasa C) y PKA (Proteína Quinasa A): Estas quinasas fosforilan sustratos que modifican la eficacia sináptica.
- Activación de CREB (proteína de unión al elemento de respuesta de AMPc): Factor de transcripción que enciende genes necesarios para la plasticidad a largo plazo.
Paso 4: Fortalecimiento sináptico — Más receptores AMPA
El resultado inmediato de la cascada de calcio es la inserción de nuevos receptores AMPA en la membrana postsináptica. Estos receptores son movilizados desde reservorios intracelulares y transportados a la superficie de la sinapsis mediante un proceso que depende de la CaMKII y la exocitosis de vesículas.
Con más receptores AMPA en la superficie, la misma cantidad de glutamato ahora produce una respuesta postsináptica más fuerte. La sinapsis se ha «potenciado». Esto constituye la LTP temprana (E-LTP), que dura de 1 a 3 horas y no requiere síntesis de nuevas proteínas.
Paso 5: Consolidación — Cambios estructurales
Si la estimulación es lo suficientemente fuerte o repetida, se activa la LTP tardía (L-LTP), que involucra:
- Transcripción génica: CREB activa genes que codifican proteínas sinápticas, factores de crecimiento como BDNF, y componentes del citoesqueleto.
- Síntesis de nuevas proteínas: Se producen proteínas que estabilizan los cambios sinápticos.
- Crecimiento de nuevas espinas dendríticas: La sinapsis no solo se fortalece — puede crecer físicamente, formando nuevos puntos de contacto.
- Remodelación del citoesqueleto: La actina filamentosa se reorganiza para estabilizar la nueva estructura sináptica.
La L-LTP puede durar semanas, meses o incluso años. Es el mecanismo que subyace a la formación de memorias duraderas.
Plasticidad en acción: evidencias del mundo real
Los taxistas de Londres: mapas cerebrales de la experiencia
Uno de los estudios más famosos sobre plasticidad cerebral fue realizado por la neurocientífica Eleanor Maguire y su equipo en el University College London, publicado en el año 2000. Los investigadores utilizaron resonancia magnética estructural para comparar el cerebro de taxistas londinenses con el de sujetos control.
Los taxistas de Londres son un grupo único: para obtener su licencia, deben aprobar «The Knowledge», un examen brutal que requiere memorizar más de 25,000 calles y miles de rutas en un radio de 10 km del centro de la ciudad. El proceso de preparación dura entre 2 y 4 años, y la tasa de aprobación es inferior al 50%.
Los resultados fueron extraordinarios: los taxistas tenían un hipocampo posterior significativamente más grande que los controles. Además, el volumen del hipocampo posterior correlacionaba positivamente con los años de experiencia como taxista. Cuanto más tiempo llevaban navegando por Londres, mayor era el cambio estructural.
Un estudio de seguimiento (Maguire et al., 2006) comparó taxistas con conductores de autobús — profesionales que también conducen todo el día pero siguen rutas fijas. Solo los taxistas, que debían navegar activamente, mostraron la hipertrofia hipocampal. Esto confirmó que el cambio no se debía simplemente a conducir, sino a la demanda cognitiva de navegación espacial.
Neurogénesis adulta
Durante décadas, el dogma neurocientífico afirmaba que nacíamos con todas las neuronas que tendríamos y que el cerebro adulto no podía generar nuevas células nerviosas. Este paradigma fue demolido en los años 1990 gracias al trabajo de Elizabeth Gould, Fred Gage y otros investigadores.
Hoy sabemos que la neurogénesis adulta ocurre en al menos dos regiones del cerebro:
- Giro dentado del hipocampo: Las nuevas neuronas aquí parecen ser cruciales para la separación de patrones — la capacidad de distinguir memorias similares (por ejemplo, dónde estacionaste hoy versus ayer).
- Zona subventricular: Las nuevas neuronas migran hacia el bulbo olfatorio, contribuyendo a la discriminación de olores.
La neurogénesis es estimulada por el ejercicio físico aeróbico, el enriquecimiento ambiental, el aprendizaje activo y factores de crecimiento como el BDNF (Factor Neurotrófico Derivado del Cerebro). Es inhibida por el estrés crónico, la privación de sueño y el aislamiento social.
Períodos críticos de plasticidad
No toda la plasticidad es igual a lo largo de la vida. Existen períodos críticos — ventanas temporales durante el desarrollo en las que el cerebro es extraordinariamente sensible a la experiencia. Los experimentos clásicos de David Hubel y Torsten Wiesel (Premio Nobel, 1981) demostraron esto con elegancia:
- Si se cierra un ojo de un gatito recién nacido durante el período crítico visual (primeras semanas de vida), las columnas de dominancia ocular en la corteza visual se reorganizan dramáticamente: las neuronas que normalmente responderían al ojo cerrado son «colonizadas» por el ojo abierto.
- Si la misma manipulación se realiza en un gato adulto, no hay efecto. El período crítico ha cerrado.
En humanos, los períodos críticos son fundamentales para la adquisición del lenguaje (hasta ~12 años para la gramática nativa), la visión binocular (primeros 2-3 años) y el vínculo afectivo. El cierre de los períodos críticos está mediado por la maduración de interneuronas GABAergicas y la formación de redes perineuronales (estructuras de matriz extracelular que estabilizan las sinapsis).
El miembro fantasma: plasticidad maladaptativa
La plasticidad no siempre es beneficiosa. Después de la amputación de un miembro, muchos pacientes experimentan sensaciones vivas — incluyendo dolor intenso — en la extremidad que ya no existe. V.S. Ramachandran demostró que esto se debe a una reorganización cortical:
En el homúnculo somatosensorial, la representación de la mano está adyacente a la de la cara. Cuando la mano es amputada, las neuronas corticales que respondían a la mano quedan «huérfanas» de input. Las fibras aferentes de la cara, vecinas en el mapa cortical, invaden el territorio desocupado. El resultado: tocar la cara del paciente evoca sensaciones en la mano fantasma. El cerebro ha «remapeado» la corteza, pero este remapeo produce una experiencia confusa y a menudo dolorosa.
Ramachandran desarrolló la ingeniosa terapia del espejo para tratar el dolor fantasma, explotando la plasticidad visual para «engañar» al cerebro y restablecer una representación coherente.
Caso clínico: Los taxistas de Londres (Maguire et al., 2000)
Eleanor Maguire y sus colegas del University College London reclutaron a 16 taxistas londinenses con una experiencia promedio de 14.3 años de conducción y los compararon con 50 sujetos control mediante resonancia magnética estructural. Los taxistas habían pasado años memorizando el complejo entramado de calles de Londres para aprobar «The Knowledge», considerado uno de los exámenes de navegación más exigentes del mundo. Las imágenes cerebrales revelaron algo notable: el hipocampo posterior de los taxistas era significativamente mayor que el de los controles, y esta diferencia aumentaba con los años de experiencia. Curiosamente, el hipocampo anterior de los taxistas era ligeramente más pequeño — sugiriendo una redistribución del tejido hipocampal, no simplemente un crecimiento global. Este hallazgo proporcionó una de las primeras demostraciones directas de que la experiencia puede esculpir la anatomía macroscópica del cerebro humano adulto.
Especialización posterior vs. anterior: El hipocampo no es una estructura homogénea. La evidencia sugiere que el hipocampo posterior está más involucrado en la representación espacial detallada — almacena «mapas cognitivos» de alta resolución del entorno. En cambio, el hipocampo anterior participa más en la codificación de nuevos eventos y el procesamiento emocional. Los taxistas necesitan mantener representaciones espaciales enormemente detalladas de miles de calles, lo que demanda específicamente los circuitos del hipocampo posterior.
Navegación activa vs. rutas fijas: Los conductores de autobús siguen rutas predeterminadas día tras día. No necesitan calcular rutas nuevas ni mantener representaciones espaciales flexibles. Su tarea se convierte en una memoria procedimental (dependiente de ganglios basales y cerebelo), no una tarea de navegación espacial activa (dependiente del hipocampo). Esto demuestra que no es la experiencia de conducir lo que cambia el hipocampo, sino la demanda cognitiva específica de navegación flexible.
Mecanismos celulares: El aumento de volumen probablemente refleja una combinación de neurogénesis en el giro dentado, aumento de la arbolización dendrítica, sinaptogénesis y posiblemente angiogénesis (formación de nuevos vasos sanguíneos). Estos cambios son mantenidos por factores neurotróficos como BDNF, cuya expresión aumenta con la actividad cognitiva intensa.
Implicación clínica: Este estudio fue revolucionario porque demostró que la plasticidad cerebral no es solo un fenómeno de laboratorio — ocurre en personas reales realizando actividades cotidianas. Abrió la puerta a la idea de que podemos «ejercitar» nuestro cerebro con demandas cognitivas apropiadas, y tiene implicaciones para la rehabilitación neurológica y la prevención del deterioro cognitivo.
Simulación interactiva: Potenciación a Largo Plazo
En esta práctica vas a simular el proceso de LTP en una sinapsis glutamatérgica. Cada vez que presiones «Estimular», la neurona presináptica liberará glutamato. Observa cómo la sinapsis responde progresivamente: primero con respuestas débiles (pocos receptores AMPA), luego con respuestas cada vez más fuertes a medida que se insertan nuevos receptores y se activan las cascadas de señalización.
Objetivo: Alcanzar una potenciación del 100% con estimulaciones repetidas. Nota cómo la frecuencia de estimulación importa — estimulaciones rápidas y consecutivas son más efectivas que estímulos aislados y espaciados.
Presiona Estimular para enviar un pulso de glutamato. Necesitarás múltiples estímulos para alcanzar el umbral de LTP.
Evaluación de dominio
Responde las siguientes preguntas para verificar tu comprensión de la plasticidad sináptica y el aprendizaje. Cada respuesta correcta suma un punto hacia tu dominio completo de esta unidad.